各種栄養素感覚受容に対する迷走神経・脊髄神経の役割解明に期待
新潟大学大学院医歯学総合研究科口腔生化学分野の市木貴子助教らの研究グループは27日、消化管における飲水後の低浸透圧感知には、肝門脈が主要な働きをしていることを明らかにしたと発表した。
マウスを用いたin vivoイメージング実験で、消化管を制御する迷走神経の感覚神経節をリアルタイムに観察し、腸管内の水による低浸透圧刺激に特異的に反応する神経群を発見した。
さらに、この低浸透圧感知には、肝門脈が消化管ホルモンを介して、主要な働きをしていることを明らかにた。これらの研究成果は、26⽇(英国時間)に英国科学誌「Nature」にオンライン公開された。
市木氏らが同研究で確立したイメージング技術は、飲水行動だけでなく、摂食行動などの別の欲求行動の制御メカニズムの解明に応用可能である。さらに、各種栄養素の感知や侵害受容といった他の感覚受容に対する迷走神経、脊髄神経の役割の解明も可能であり、さらなる研究の発展が期待される。
飲水量を調整し、体液恒常性を維持することは、生物が生きる上で必要不可欠だ。これまで、飲水後に消化管内で浸透圧変化が感知されることで、脳内の飲水促進ニューロンが抑制され、飲水抑制が起こることが示唆されてきたが、そのメカニズムは長らく不明であった。
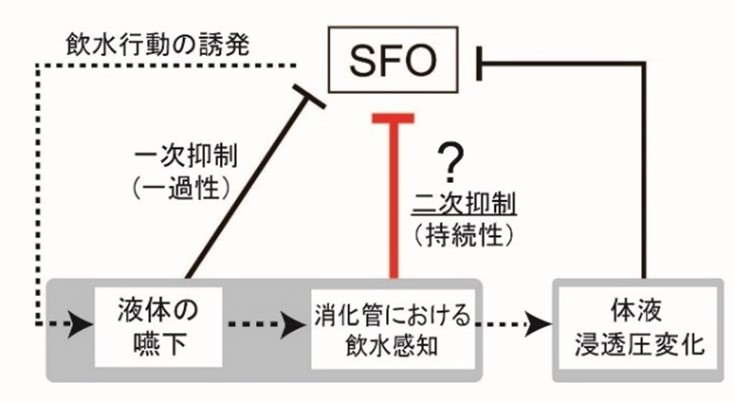
適切な飲水欲求の調節は、体液恒常性の維持に必要不可欠である。神経系による飲水制御メカニズムとして、脳内の脳弓下器官(SFO, subfornical organ)に存在する神経群が飲水促進に寄与する。
このSFOの飲水促進ニューロンは、飲水後に体液浸透圧が変化する以前に強く抑制されるため、神経系による飲水抑制機構の存在が示唆されてきた。飲水後の飲水抑制には、液体嚥下に反応して起こる一次抑制と、消化管での浸透圧感知によって持続性に働く二次抑制があると考えられている(図1)。
二次抑制のメカニズムには、不明な点が多く残されており、消化管における飲水感知シグナルが中枢神経系に伝達される仕組みは分かっていなかった。
市木氏らは、確立したin vivoイメージングの実験系を用いて、消化管内の浸透圧変化がどのように感知され、飲水後の飲水抑制に寄与するのかを調べた。
消化管の感覚受容には、迷走神経が重要な役割を果たしている。これまでに、消化管内の浸透圧変化に対する迷走神経の応答は部分的な知見しか得られておらず、飲水に応じた低浸透圧刺激が迷走神経を活性化するかは不明であった。
まず、同研究グループは、マウスの迷走神経群から電気生理学的手法を用いて神経活動を記録する実験系を構築した。具体的には、マウスin vivoにおいて飲水を模した腸管内への水灌流刺激を行うと同時に、迷走神経の応答を複合活動電位として記録する手法を確立。
このin vivo複合活動電位記録の結果、迷走神経が腸管内の水刺激に強く応答することを見出した。
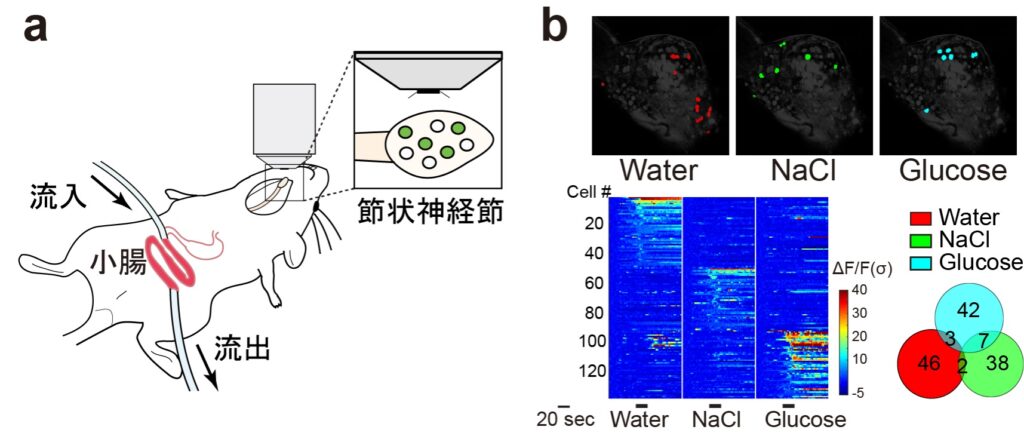
次に、in vivoカルシウムイメージングによる単一細胞レベルでの神経応答の可視化を行うため、迷走神経の求心性感覚神経節である節状神経節のin vivoイメージングの実験系を確立した(図2a)。
具体的には、感覚神経依存的にカルシウムセンサー(GCaMP)を発現するマウスを用いて、腸管内への水灌流刺激を行うと同時に、節状神経節ニューロンの応答を観察した。
その結果、迷走神経において、水による低浸透圧刺激に特異的に応答する神経群の存在が明らかになった(図2b)。
一方で、消化管の感覚受容は迷走神経のみならず、脊髄神経の求心性感覚神経も担っており、脊髄神経も飲水抑制に何らかの役割を果たす可能性がある。その可能性を考慮し、脊髄後根神経節のin vivoイメージングの実験系を確立し、反応を観察した。
その結果、皮膚への触刺激や酢酸溶液による侵害刺激に対する反応が観察された一方で、腸管への水灌流刺激に対してはほとんど応答が見られなかった。これにより、迷走神経と比較して、脊髄神経は腸管での低浸透圧感知にはほとんど寄与しないことが示唆された。
次に、消化管内の低浸透圧に反応する神経群のマーカー遺伝子を探索し、低浸透圧応答神経群を含む遺伝子tachykinin 1 (Tac1)と、含まない遺伝子としてneurotensin (Nts)を同定した。それぞれの遺伝子のプロモーターでドライブされるCreマウスに、Cre依存的に蛍光色素を遺伝子導入するアデノ随伴ウィルスを節状神経節にインジェクションし、末梢器官への投射分布を確認した。
その結果、Tac1-Creで消化管と肝臓を繋ぐ血管である肝門脈への投射を確認したのに対し、Nts-Creでは認めらなかったため、肝門脈が低浸透圧感知に寄与するのではないかと考えた。
腸管から吸収された水分、栄養素は、上腸間膜静脈を経由してすべて肝門脈へ集められるため、肝門脈を支配する神経群が、水分や栄養素の感知に何らかの役割を果たす可能性が示唆された。
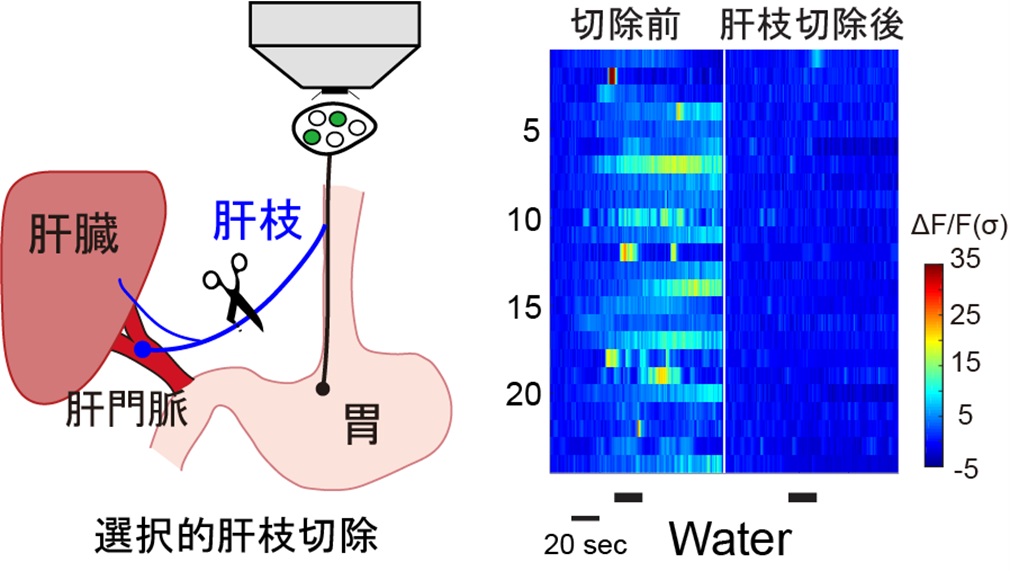
そこで、肝門脈を支配する迷走神経の分枝(肝枝)を切除し、腸管への低浸透圧刺激に対する応答を調べた。その結果、切除後に低浸透圧刺激に対する応答が消失することを確認した(図3)。
マウス行動実験においても、肝枝切除後に脱水後の飲水量が有意に増加することが確認された。さらに、脳内のSFOの飲水時のリアルタイム神経活動をファイバーフォトメトリによって観察したところ、肝枝切除後に、飲水後のSFO飲水促進ニューロンの活動抑制が阻害されることが確認された。
これらの結果から、末梢器官において、肝門脈を支配する迷走神経が低浸透圧刺激の感知に寄与することが明らかとなった。
次に、肝門脈が直接的に低浸透圧刺激を感知するかどうかを調べるために、肝門脈へカテーテルを挿入し、低張食塩水を注入した際に、迷走神経の応答が見られるか調べた。その結果、肝門脈への直接的な低浸透圧刺激に対する応答は認められなかった。
そこで、消化管ホルモンの関与を想定し、主要な消化管ホルモンを肝門脈へ注入した際の迷走神経応答を調べた。各種ホルモンのうち、血管作動性腸管ペプチド(VIP, vasoactive intestinal peptide)による迷走神経応答が、肝枝切除後に消失し、かつ、腸管の水灌流によって応答する神経群と4割程度同一であることを見出した。
これにより、飲水後に腸管にて低浸透圧が感知された後、VIPをはじめとする消化管ホルモンが分泌され、肝門脈に作用することで、迷走神経を介して中枢に飲水抑制シグナルが送られている可能性が示唆された。
同研究により、長らく不明であった消化管における飲水抑制回路が明らかとなり、生物が生きる上で必須な体液恒常性を維持する神経基盤の一端が解明されました。
今回、消化管への低浸透圧刺激に対し、VIPをはじめとする消化管ホルモンが肝門脈に作用することで、迷走神経応答が起きていることが示唆されたが、腸管においてどのようにして低浸透圧が感知され、ホルモン分泌が行われているのかは明らかになっていない。
また、迷走神経の節状神経節からのシグナルがどういった経路を介して脳内のSFOニューロンに伝達されているのかに関してもよくわかっていない。
今後、市木氏らは、これらを明らかにしていく意向を示している。

