自己炎症性・神経変性疾患の新規治療開発への応用に期待
東北大学大学院生命科学研究科の田口友彦教授および岐阜大学糖鎖生命コア研究所の鈴木健一教授らの共同研究グループは、ヘルペスウイルスなどのDNAウイルスの感染に応答して自然免疫応答・炎症を誘導するSTINGタンパク質の活性化機構を完全解明した。
同研究成果により細胞内コレステロールレベルを制御することが自己炎症性・神経変性疾患を治療する新しい手段になると期待される。
なお、これらの研究成果は、2024年1月11日に科学誌Nature Communicationsに掲載された。
自然免疫は、先天的に備わってい、異物に対する応答機構だ。STINGタンパク質は、ヘルペスウイルスなどのDNAウイルスの感染に応答して自然免疫応答、炎症を誘導する。それによってSTING経路はDNAウイルス感染から身体を守っているが、その一方で、異常な活性化は自己免疫疾患、神経変性疾患、がんなど多様な疾患を引き起こす。
これまで、同共同研究グループは、STINGの活性にはパルミトイル化脂質修飾が必要であることを明らかにしてきた。今回、STINGは、パルミトイル化脂質修飾を介してTGNのコレステロールを含む脂質マイクロドメインで平均20分子以上のクラスターを形成し、STINGのクラスター形成が自然免疫シグナルの活性化に必要であることを明らかにした。
同研究成果により細胞内コレステロールレベルを制御することがこれら疾患を治療する新しい手段になると期待される。
自然免疫は、体内に侵入した病原体や異常になった自己の細胞をいち早く感知し、それを排除する仕組みであり、生体防御の最前線に位置している。小胞体に局在する4回膜貫通タンパク質STINGは、DNAウイルスの感染により細胞質に出現したDNAに応答し、自然免疫応答を誘導する。
最近、STING経路は、がん、自己炎症性疾患、老化性炎症、パーキンソン病、ハンチントン病、筋萎縮性側索硬化症など多様な疾患の炎症病態に寄与することが報告され、非常に注目されている。
これまでに東北大学大学院生命科学研究科の向井助教・田口教授のグループは、DNAウイルス感染に伴ってSTINGが小胞体からゴルジ体へ移動することを見出した。さらに、ゴルジ体でSTINGがパルミトイル化脂質修飾を受けることは、自然免疫応答の活性化に必要であることを明らかにしてきた(Mukai et al. Nat Commun 2016)。
だが、パルミトイル化STINGがどのように自然免疫応答を活性化するのかは不明であった。そこで、同研究グループは、まず、生細胞内におけるSTINGクラスターの存在を検証した。
パルミトイル化脂質修飾を受けたタンパク質は、コレステロールに富む脂質マイクロドメイン内でクラスターを形成して機能を発揮すると考えらるため、脂質マイクロドメイン内でパルミトイル化STINGがクラスター化することで、自然免疫応答を活性化するという仮説を立てた。
同研究では、1分子レベルで細胞内分子の局在を観察できる超解像顕微鏡法PALMを用いて、STING分子の細胞内での動態を解析した。
その結果、STINGはTGNにおいて平均20分子以上のクラスターを形成することが判明した(図2)。
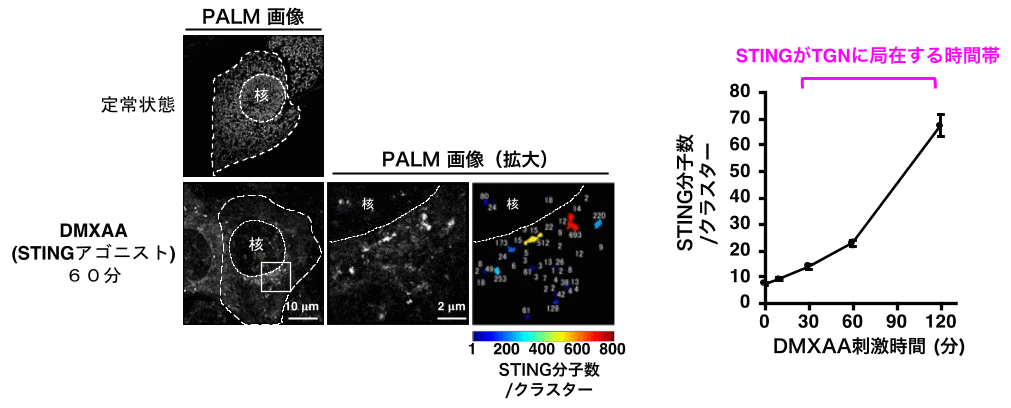
STINGは定常状態において、細胞質全体に網目状に分布する小胞体に局在している。。DNAウイルスの感染等により刺激されるとSTINGは核近辺のゴルジ体に移動する。この際、PALMによりSTINGのクラスター形成を定量的に評価すると、STINGがTGNに局在する時間帯において、STINGクラスター内に含まれるSTING分子数が急激に上昇した。
続いて、STINGのクラスター形成を制御する分子機構の解明に挑んだ。まず、パルミトイル化脂質修飾がSTINGのクラスター形成に寄与するのか検討した。
STING特異的パルミトイル化阻害剤を用いて、STINGのパルミトイル化を阻害したところ、STINGのクラスター形成が有意に抑制された(図3)。
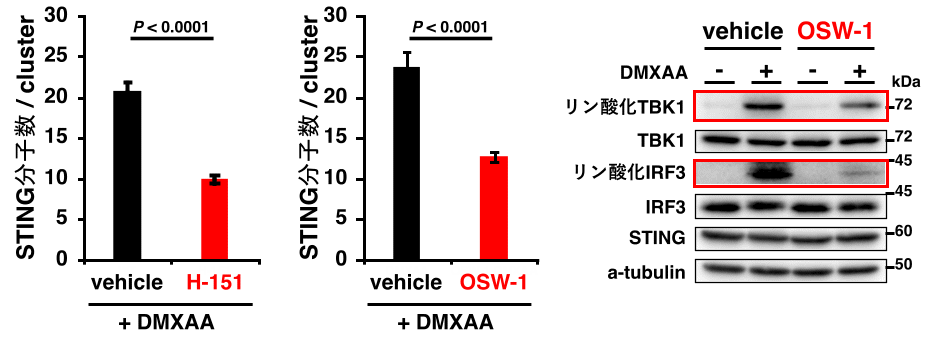
STING特異的パルミトイル化阻害剤H-151を処理すると、STINGのクラスター形成が抑制された。OSW-1によりTGNへのコレステロール輸送を阻害すると、STINGのクラスター化が抑制された。さらに、OSW-1処理により、STING経路が自然免疫シグナルを活性化するために必要なTBK1およびIRF3のリン酸化が抑制された。
次に、STINGのクラスター形成における脂質マイクロドメインの必要性を検討した。TGNへのコレステロール輸送を阻害したところ、STINGのクラスター形成が有意に抑制された。
この際、STING経路が自然免疫応答を活性化するために必要なキナーゼTBK1および転写因子IRF3のリン酸化も抑えられた(図3)。
以上、STINGのクラスター形成はSTINGのパルミトイル脂質修飾およびコレステロールに依存していることが示された。
STINGが自然免疫シグナルを活性化するためには、リン酸化酵素であるTBK1がSTING上に安定的に滞在し、STINGタンパク質などをリン酸化する必要がある。そこで、STINGのクラスター形成が、TBK1のSTING上での滞在時間に影響するのかを検証した。
その結果、STINGのクラスターが形成されない条件下では、TBK11分子あたりのSTING上での滞在時間が減少することがわかった。
約四半世紀前、コレステロールに富む脂質マイクロドメインにパルミトイル化脂質修飾を受けたタンパク質が集積することでその機能を発揮するモデルが提唱された(脂質ラフト仮説、Simons et al. Nature 1997)。
だが、脂質ラフトに局在すると考えられているタンパク質を一分子レベルで観察することは難しく、これまで脂質ラフトの存在証明や機能解析は進んでいなかった。
今回、PALMを用いることで、STINGタンパク質がTGN上でクラスター(20分子以上)を形成することを発見した。さらに、STINGのパルミトイル化脂質修飾やTGNへのコレステロール輸送を阻害すると、STINGクラスターが形成できず、自然免疫応答も顕著に抑制されることがわかった。
この結果は、パルミトイル化STINGがTGNのコレステロールに富む脂質ラフト上で集積し、その機能を発揮することを示している。従って、同研究成果は、細胞内における脂質ラフトの存在とその機能を直接的に証明するものであり、基礎生物学的に重要な発見である。(図1)。
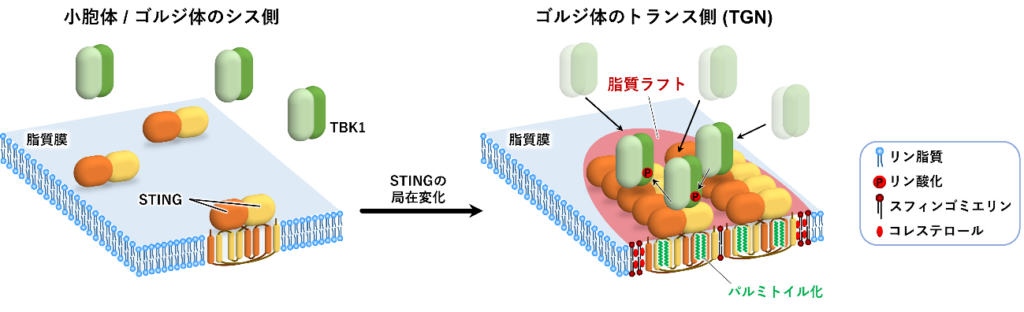
小胞体もしくはゴルジ体のシス側に局在するSTINGはクラスターを形成しておらず、TBK1は細胞質に局在している。一方、TGNに移行したSTINGはパルミトイル化脂質修飾依存的に脂質ラフト上でクラスターを形成する。その結果、TBK1が安定的にSTING上に局在できるようになり、TBK1が活性化する。活性化したTBK1がSTINGなどをリン参加することで自然免疫シグナルを活性化する。
STINGはウイルス由来の非自己DNAに応答して、自然免疫シグナルを誘導するタンパク質として発見された。最近STING自然免疫応答は、がん免疫応答において重要な役割を果たすことや、老化、自己炎症性疾患、および神経変性疾患における炎症病態に寄与することが報告され、疾患治療のターゲットとして非常に注目されている。
今回、TGNへのコレステロール輸送を阻害するとSTING自然免疫応答が顕著に抑制されることを示した。従って、同研究成果から、コレステロールレベルを減らすことは、STING炎症と関連する上記疾患を治療する新しい手段となることが期待される。

